Natural killer cells lull tumours into dormancy
Natural killer cells can drive spreading cancer cells to enter a state of dormancy. That finding, together with the discovery of a pathway that hinders this antitumour function, could spur the development of new treatments.
Noella Lopes & Eric Vivier
Efforts to treat tumours that have spread from their initial site in the body to grow elsewhere are often unsuccessful. Such tumours, called metastases, are the main cause of cancer-related deaths, so finding a way to control them is crucial to meeting this medical need. Before metastases begin to grow, cancer cells might have already migrated from the primary tumour to seed various other sites (a process called metastasis), where they can remain dormant for long periods of time. Surveillance by immune cells is known to help to maintain this dormancy1, but the mechanisms involved in the switch from dormancy to the growth of metastases have been unclear — until now. Writing in Nature, Correia et al.2 report the pivotal role of natural killer (NK) cells in controlling the development of liver metastases arising from breast cancer.
NK cells are part of the innate branch of the immune system. They can kill other cells and produce soluble messenger molecules, called cytokines and chemokines, that regulate immune responses3. The ability of NK cells to detect and eliminate a wide array of tumour cells directly, and their capacity to shape antitumour immune responses by making cytokines or chemokines, have led to the development of clinical strategies that harness their anticancer functions3–5.
Several studies have suggested that NK cells specialize in eliminating metastases rather than targeting tumour cells at their primary site of growth 6. For some cancers, people who have more tumour-infiltrating NK cells seem to have fewer metastases, as seen in those with cancers such as gastrointestinal sarcoma, and gastric, colorectal, renal or prostate carcinoma3,6. The depletion or dysfunction of NK cells in mice also results in an increase in metastases3. By contrast, when their normal regulation is removed, NK cells protect against the spread of tumours to the liver and lungs7. Tumour cells entering dormancy downregulate their expression of ligand molecules that can activate NK cell receptors, and become resistant to killing mediated by NK cells8.
Correia and colleagues decided to further investigate the composition and dynamics of tumour cells in dormancy. One approach they took was to study the gene-expression profile of human and mouse breast cancer cells transplanted into mice. These cells underwent metastasis to reach sites such as the liver, where they became dormant tumour cells. The authors assessed genes expressed by cells in the vicinity of the dormant tumour cells in the surrounding stromal tissue. These data revealed a gene signature associated with responses mediated by NK cells. Furthermore, Correia et al. compared the areas around dormant tumour cells with those in tumour-free livers, and found that NK cells were the only type of immune cell to increase in number during dormancy. This suggests that NK cells have a crucial role in events that block the reawakening of dormant tumour cells (Fig. 1).
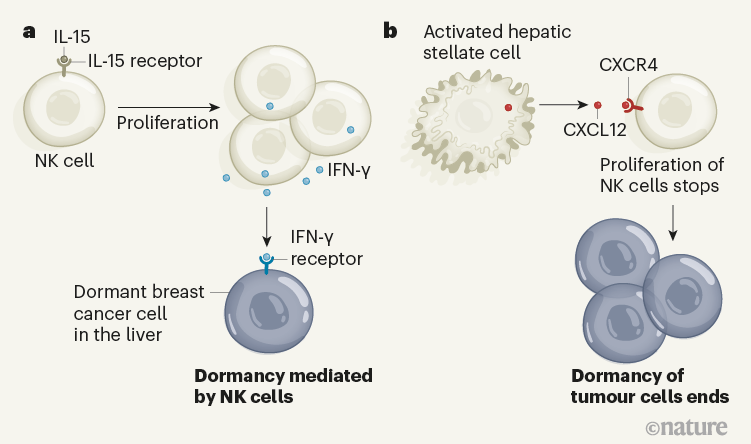 Figure 1 | Interactions that affect the dormancy of tumour cells. Correia et al.2 report mouse and human evidence indicating that a type of immune cell called a natural killer (NK) cell has a key role in preventing the growth of spreading tumour cells, called metastases. a, The molecule IL-15 drives NK cells to proliferate, and these cells secrete the protein IFN- γ, which keeps breast cancer cells that have migrated to the liver in a dormant state. b, Exit from this state of dormancy is associated with activated hepatic stellate cells, which secrete the molecule CXCL12. CXCL12 binds to the CXCR4 receptor on NK cells. This results in NK cells ceasing to divide and stops them from promoting tumour dormancy. The tumour cells proliferate as a consequence.
Figure 1 | Interactions that affect the dormancy of tumour cells. Correia et al.2 report mouse and human evidence indicating that a type of immune cell called a natural killer (NK) cell has a key role in preventing the growth of spreading tumour cells, called metastases. a, The molecule IL-15 drives NK cells to proliferate, and these cells secrete the protein IFN- γ, which keeps breast cancer cells that have migrated to the liver in a dormant state. b, Exit from this state of dormancy is associated with activated hepatic stellate cells, which secrete the molecule CXCL12. CXCL12 binds to the CXCR4 receptor on NK cells. This results in NK cells ceasing to divide and stops them from promoting tumour dormancy. The tumour cells proliferate as a consequence.
Consistent with this hypothesis, the authors report that depleting NK cells in a mouse tumour model then led to higher levels of metastases in the liver. However, if NK cells were boosted using the cytokine IL-15, this prevented the formation of liver metastases and tumour cells remained dormant. The authors’ results demonstrate that the size of the pool of NK cells in the liver environment determines whether dormancy occurs or metastases form.
The liver environment associated with dormant tumour cells contained NK cells producing the cytokine interferon-γ (IFN-γ). Correia and colleagues report that, in vitro, adding IFN-γ can nudge cancer cells into dormancy — consistent with the idea that IFN-γ has a key role in controlling the cancer dormancy mediated by NK cells.
Might other factors disrupt NK cells and thereby promote the formation of metastases? A clue to this came from the authors’ discovery that a pool of activated hepatic stellate cells found in the mouse liver increased when tumours switched from dormancy to forming metastases. Hepatic stellate cells have been identified as the main disease-driving population of cells for a condition called hepatic fibrosis9, in which the liver becomes damaged and scarred. These changes often precede tumour formation. The accumulation of activated hepatic stellate cells occurs at the same time as a decline in NK cells, owing to a decrease in NK-cell proliferation. The authors’ results suggest that activated hepatic stellate cells promote metastases in the liver by inhibiting NK cells, thereby disrupting cancer dormancy.
Correia et al. found that hepatic stellate cells secrete the chemokine CXCL12, which has been implicated in aiding the directional migration of breast cancer cells10. Organs that express the highest levels of CXCL12 are the most common sites of metastasis in human breast cancer10. Human NK cells in the liver have a receptor, called CXCR4, that recognizes CXCL12. Correia and colleagues report that activated hepatic stellate cells hamper the function of NK cells in the liver through CXCL12–CXCR4 interactions that halt the proliferation of NK cells, thereby tipping the scales from tumour dormancy to the promotion of metastasis. This study thus reveals a previously unknown function of CXCL12 in altering NK-cell-mediated immunity, in addition to its known effects on tumour cells 10.
The authors next examined pairs of human biopsy specimens from metastases and healthy adjacent liver tissue, taken from people with breast cancer. Consistent with the data from mice, the analysis showed that activated hepatic stellate cells accumulated in metastases, and that their abundance was inversely correlated with that of NK cells. The authors’ analysis of published gene-expression data for colorectal cancer that has metastasized to the liver revealed the same association, suggesting that this cellular crosstalk might be relevant for the growth of other types of spreading cancer.
Several questions remain to be answered. For example, the mechanisms underlying the accumulation of NK cells associated with dormant tumour cells and the triggering of IFN-γ production in these circumstances remain to be fully determined. It is not completely clear how CXCL12 that is secreted by activated hepatic stellate cells hinders the function of NK cells. Furthermore, determining whether the CXCL12–CXRC4 axis awakens dormant tumour cells in humans is of utmost importance, and, if so, in which types of cancer.
Finally, the similarities between NK cells and another sort of immune cell called type 1 innate lymphoid cells (ILC1) should prompt further investigation of the role of ILC1 in controlling metastasis3. Indeed, these cells have a complex role in tumour responses11,12. Correia et al. excluded ILC1 as having a role in controlling metastases, because they observed no notable changes in the level of these cells when comparing tumour dormancy and metastases in the liver. However, the lack of a specific ILC1-deficient mouse model means that it is not possible to precisely dissect the respective roles of NK cells and ILC1 in the control of metastasis, leaving a key question unresolved.
By showing that the IFN-γ-driven effects of NK cells maintain breast cancer cells in a dormant state, Correia and colleagues have revealed that NK cells have other and previously unsuspected anticancer capacities. This finding paves the way for the development of cancer treatment strategies that prevent dormant reservoirs of tumour cells from awakening. For instance, molecules that strongly stimulate the IL-15 pathway in NK cells are already available. These IL-15 superagonists, such as ALT-803 or NKTR-255, are being tested in clinical trials3,5, and the rationale for their use should now also take into account the role of NK cells in controlling dormant tumour cells.
Furthermore, drugs that inhibit CXCR4 are being developed. It would be interesting to determine whether these inhibitors could help to sustain the activity of NK cells in maintaining tumour dormancy. In addition, engineered antibodies called NK cell engagers, which can stimulate NK cells and form a bridge that connects them to tumour cells, offers another way to promote the function of NK cells13. Current clinical trials are also testing various approaches to manipulate NK cells for therapeutic benefit3–5. Besides the well-characterized effects of NK cells in tumour immunity, Correia and colleagues’ work further highlights the possible advantages of harnessing NK cells to target cancers.
doi: https://doi.org/10.1038/d41586-021-01381-5
https://www.nature.com/articles/d41586-021-01381-5
- Mohme, M., Riethdorf, S. & Pantel, K. Nature Rev. Clin. Oncol. 14, 155–167 (2017).
- Correia, A. et al. Nature https://doi.org/10.1038/s41586- 021-03614-z (2021).
- Chiossone, L., Dumas, P.-Y., Vienne, M. & Vivier, E. Nature Rev. Immunol. 18, 671–688 (2018).
- Daher, M. & Rezvani, K. Cancer Discov. 11, 45–58 (2021).
- Myers, J. A. & Miller, J. S. Nature Rev. Clin. Oncol. 18, 85–100 (2021).
- López-Soto, A., Gonzalez, S., Smyth, M. J. & Galluzzi, L. Cancer Cell 32, 135–154 (2017).
- Molgora, M. et al. Nature 551, 110–114 (2017).
- Malladi, S. et al. Cell 165, 45–60 (2016).
- Tsuchida, T. & Friedman, S. L. Nature Rev. Gastroenterol. Hepatol. 14, 397–411 (2017).
- Shi, Y., Riese, D. J., 2nd & Shen, J. Front. Pharmacol. 11, 574667 (2020). 11. Dadi, S. et al. Cell 164, 365–377 (2016).
- Gao, Y. et al. Nature Immunol. 18, 1004–1015 (2017).
- Gauthier, L. et al. Cell. 177, 1701–1713 (2019).